
Analysis of Height Growth Suggests Moderate Growth of Tilia cordata and Acer platanoides at the Native Hemiboreal Stands in Latvia
 ,
, 
Abstract
:1. Introduction
2. Material and Methods
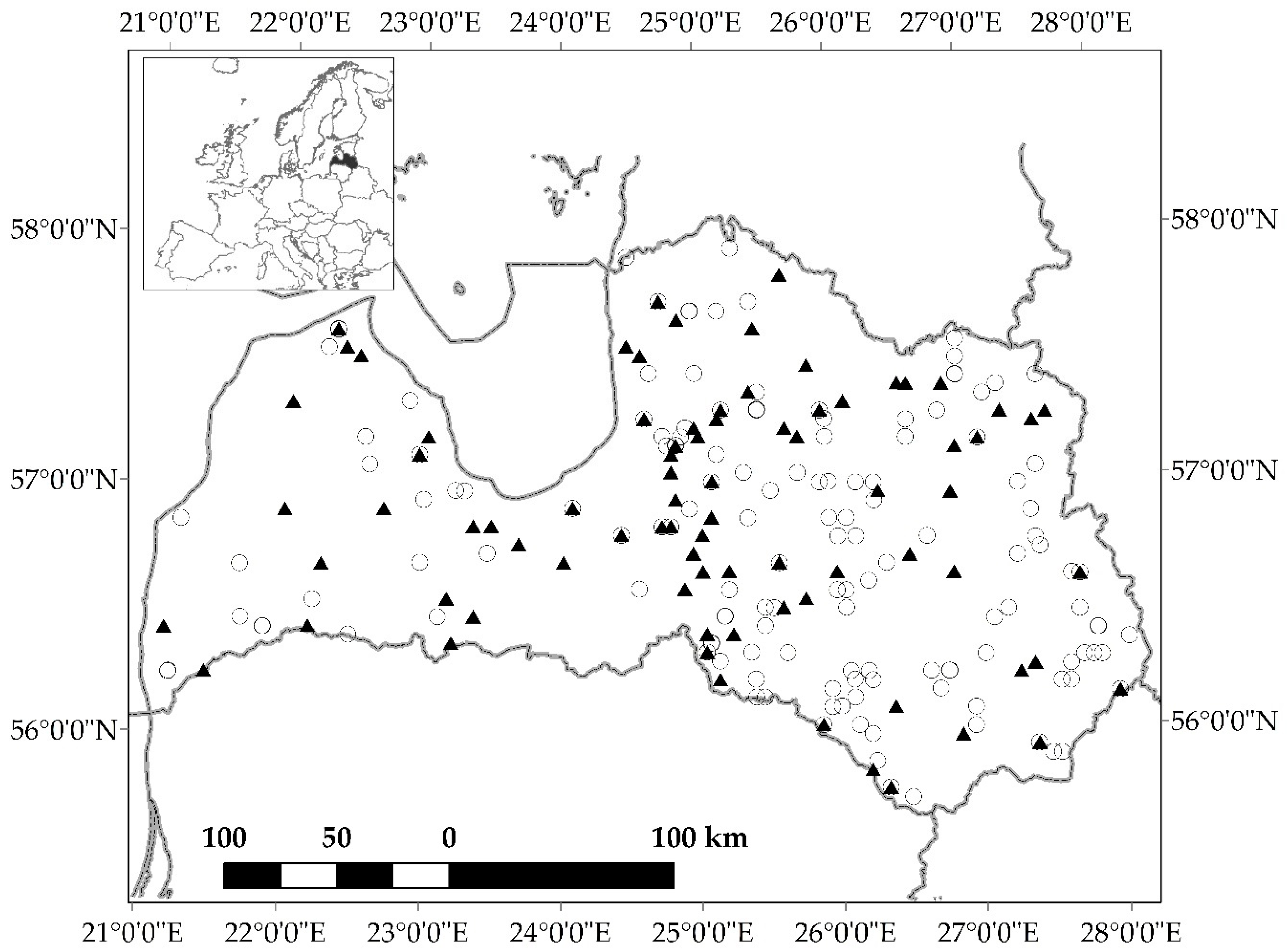
2.1. Study Area and Measurements
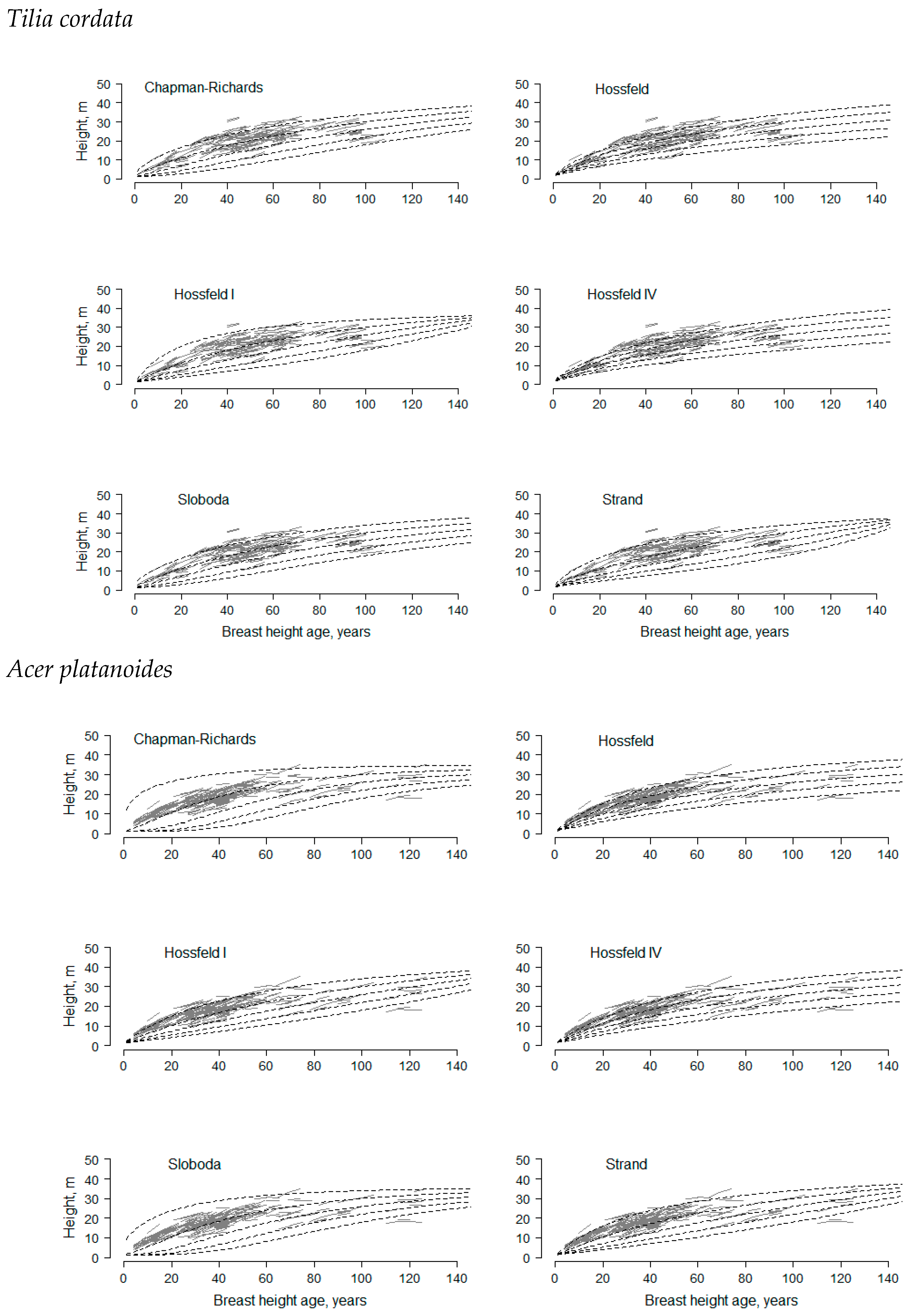
2.2. Model Fitting

{kind=link}
{kind=link}
{kind=link}
| Base Model | Generalised Algebraic Difference Approach Model | |
|---|---|---|
| Chapman–Richards: | with and | [47] |
| Hossfeld | [30] | |
| Hossfeld I | [34] | |
| Hossfeld IV | [47] | |
| Sloboda | [48] | |
| Strand | [34] |
3. Results
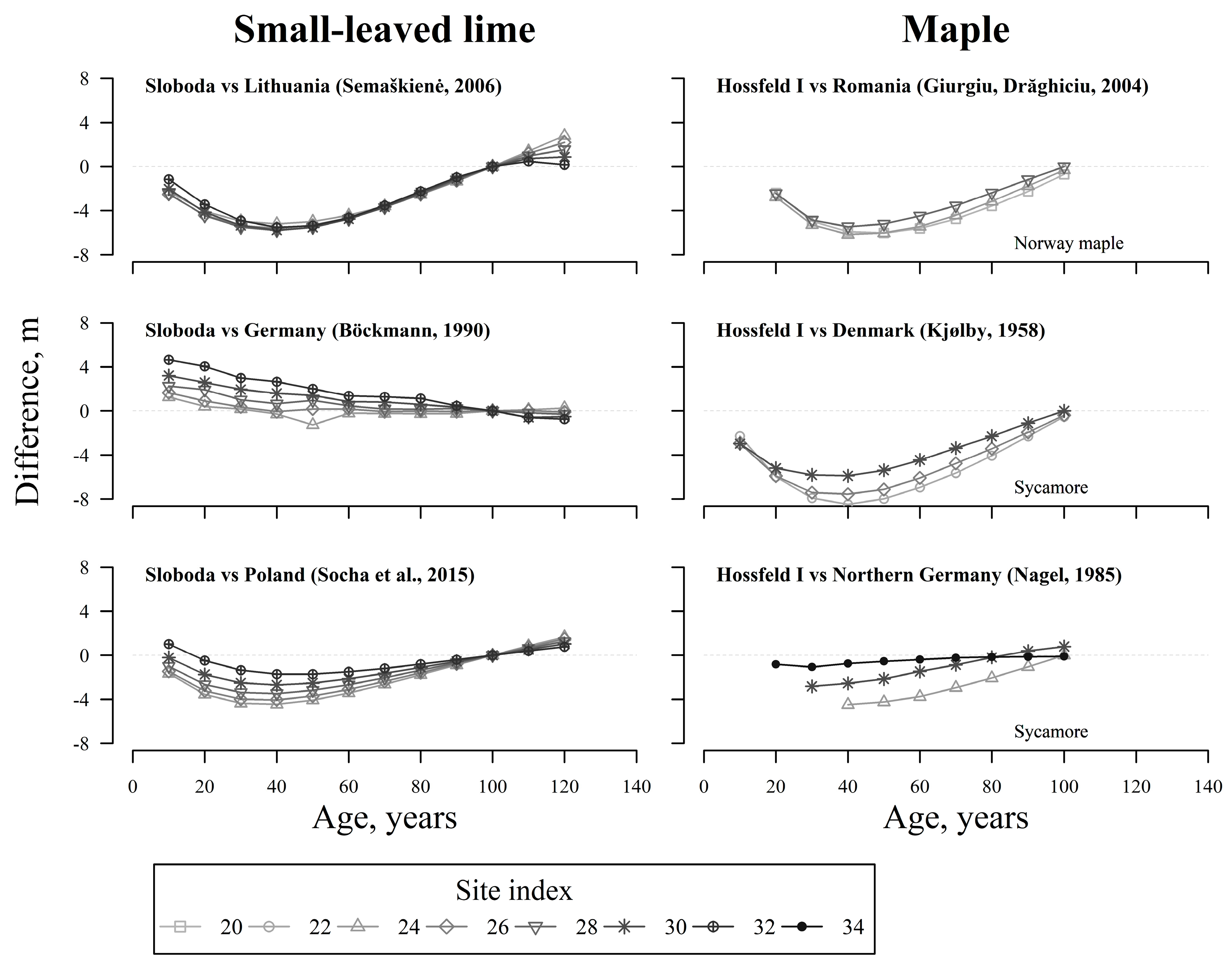
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Huuskonen, S.; Domisch, T.; Finér, L.; Hantula, J.; Hynynen, J.; Matala, J.; Miina, J.; Neuvonen, S.; Nevalainen, S.; Niemistö, P.; et al. What is the potential for replacing monocultures with mixed-species stands to enhance ecosystem services in boreal forests in Fennoscandia? For. Ecol. Manag. 2021, 479, 118558. [Google Scholar] [CrossRef]
- Huuskonen, S.; Lahtinen, T.; Miina, J.; Uotila, K.; Bianchi, S.; Niemistö, P. Growth Dynamics of Young Mixed Norway Spruce and Birch Stands in Finland. Forests 2022, 14, 56. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y. Interactions between overstory and understorey vegetation along an overstorey compositional gradient. J. Veg. Sci. 2013, 24, 543–552. [Google Scholar] [CrossRef]
- Grossiord, C. Having the right neighbors: How tree species diversity modulates drought impacts on forests. New Phytol. 2018, 228, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Buras, A.; Menzel, A. Projecting tree species composition changes of European forests for 2061–2090 under RCP 4.5 and RCP 8.5 scenarios. Front. Plant Sci. 2019, 9, 1986. [Google Scholar] [CrossRef] [PubMed]
- Götmark, F.; Fridman, J.; Kempe, G.; Norden, B. Broadleaved tree species in conifer-dominated forestry: Regeneration and limitation of saplings in southern Sweden. For. Ecol. Manag. 2005, 214, 142–157. [Google Scholar] [CrossRef]
- Laiviņš, M.; Čekstere, G.; Kaupe, D. Nemoralisation of Pinus sylvestris and Picea abies forest stands in the hemi-boreal zone: A case study from Latvia. Proc. Latv. Acad. Sci. 2021, 75, 299–309. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Koufaki, T.; Sazeides, C.I.; Spyroglou, G.; Theodorou, K. Potential impacts of climate change on the habitat suitability of the dominant tree species in Greece. Plants 2022, 11, 1616. [Google Scholar] [CrossRef]
- Daugaviete, M.; Lazdina, D.; Celma, S.; Daugavietis, U. Characteristic growing parameters of small-leaved lime and Norway maple stands in the climatic conditions of Latvia. Acta Biol. Univ. Daugav. 2019, 19, 115–128. [Google Scholar]
- Radoglou, K.; Dobrowolska, D.; Spyroglou, G.; Nicolescu, V.N. A review on the ecology and silviculture of limes (Tilia cordata Mill., Tilia platyphyllos Scop. and Tilia tomentosa Moench.) in Europe. Die Bodenkultur 2009, 60, 9–20. [Google Scholar]
- Caudullo, G.; de Rigo, D. Acer platanoides in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publications Office of the EU: Luxembourg, 2016; p. e019159. [Google Scholar]
- Eaton, E.; Caudullo, G.; De Rigo, D. Tilia cordata, Tilia platyphyllos and other limes in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the EU: Luxembourg, 2016; pp. 184–185. [Google Scholar]
- Ruņģis, D.E.; Krivmane, B. Assessment of the Structure and Diversity of Latvian Acer platanoides Populations Using Cross-Species Nuclear Microsatellite Markers. Proc. Latv. Acad. Sci. 2021, 75, 254–260. [Google Scholar] [CrossRef]
- Ruņģis, D.E.; Krivmane, B. Genetic diversity and structure of Latvian Tilia cordata populations. Proc. Latv. Acad. Sci. 2021, 75, 261–267. [Google Scholar] [CrossRef]
- Carón, M.M.; De Frenne, P.; Brunet, J.; Chabrerie, O.; Cousins, S.A.O.; De Backer, L.; Decocq, G.; Diekmann, M.; Heinken, T.; Kolb, A.; et al. Interacting effects of warming and drought on regeneration and early growth of Acer pseudoplatanus and A. platanoides. Plant Biol. 2015, 17, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Kunz, J.; Löffler, G.; Bauhus, J. Minor European broadleaved tree species are more drought-tolerant than Fagus sylvatica but not more tolerant than Quercus petraea. For. Ecol. Manag. 2018, 414, 15–27. [Google Scholar] [CrossRef]
- Zimmermann, J.; Hauck, M.; Dulamsuren, C.; Leuschner, C. Climate warming-related growth decline affects Fagus sylvatica, but not other broad-leaved tree species in Central European mixed forests. Ecosystems 2015, 18, 560–572. [Google Scholar] [CrossRef]
- Bosco, C.; de Rigo, D.; Dewitte, O.; Poesen, J.; Panagos, P. Modelling soil erosion at European scale: Towards harmonization and reproducibility. Nat. Hazard. Earth. Sys. 2015, 15, 225–245. [Google Scholar] [CrossRef]
- Hytteborn, H.; Maslov, A.A.; Nazimova, D.I.; Rysin, L.P. Coniferous Forests. In Ecosystems of the World; Andersson, F.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 6, pp. 23–99. [Google Scholar]
- De Jaegere, T.; Hein, S.; Claessens, H. A review of the characteristics of small-leaved lime (Tilia cordata Mill.) and their implications for silviculture in a changing climate. Forests 2016, 7, 56. [Google Scholar] [CrossRef]
- Hemery, G.E.; Clark, J.R.; Aldinger, E.; Claessens, H.; Malvolti, M.E.; O’connor, E.; Raftoyannis, Y.; Savill, P.S.; Brus, R. Growing scattered broadleaved tree species in Europe in a changing climate: A review of risks and opportunities. Forestry 2010, 83, 65–81. [Google Scholar] [CrossRef]
- Lazic, D.; George, J.P.; Rusanen, M.; Ballian, D.; Pfattner, S.; Konrad, H. Population Differentiation in Acer platanoides L. at the Regional Scale—Laying the Basis for Effective Conservation of Its Genetic Resources in Austria. Forests 2022, 13, 552. [Google Scholar] [CrossRef]
- Matisone, I.; Matisons, R.; Jansons, A. The Struggle of Ash—Insights from Long-Term Survey in Latvia. Forests 2021, 12, 340. [Google Scholar] [CrossRef]
- Pigott, D. Lime-trees and Basswoods: A Biological Monograph of the Genus Tilia, 1st ed.; Cambridge University Press: New York, NY, USA, 2012; p. 405. [Google Scholar]
- Mikola, J.; Koikkalainen, K.; Rasehorn, M.; Silfver, T.; Paaso, U.; Rousi, M. Genotypic traits and tradeoffs of fast growth in silver birch, a pioneer tree. Oecologia 2021, 196, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-González, J.G.; Ruiz-González, A.D.; Rodríguez-Soalleiro, R.; Barrio-Anta, M. Ecorregional site index models for Pinus pinaster in Galicia (northwestern Spain). Ann. For. Sci. 2005, 62, 115–127. [Google Scholar] [CrossRef]
- Socha, J.; Tymińska-Czabańska, L.; Bronisz, K.; Zięba, S.; Hawryło, P. Regional height growth models for Scots pine in Poland. Sci. Rep. 2021, 11, 10330. [Google Scholar] [CrossRef] [PubMed]
- Cieszewski, C.J.; Bailey, R.L. Generalized algebraic difference approach: Theory based derivations with polymorphism and variable asymptotes. For. Sci. 2000, 46, 115–126. [Google Scholar] [CrossRef]
- Hipler, S.M.; Spiecker, H.; Wu, S. Dynamic Top Height Growth Models for Eight Native Tree Species in a Cool-Temperate Region in Northeast China. Forests 2021, 12, 965. [Google Scholar] [CrossRef]
- Cieszewski, C.J. Comparing fixed- and variable-base-age site equations having single versus multiple asymptotes. Forest Sci. 2002, 48, 7–23. [Google Scholar] [CrossRef]
- Cieszewski, C.J. Three methods of deriving advanced dynamic site equations demonstrated on inland Douglas-fir site curves. Can. J. Forest Res. 2001, 31, 165–173. [Google Scholar] [CrossRef]
- Cieszewski, C.J.; Strub, M.; Zasada, M. New dynamic site equation that fits best the Schwappach data for Scots pine (Pinus sylvestris L.) in Central Europe. For. Ecol. Manag. 2007, 243, 83–93. [Google Scholar] [CrossRef]
- Manso, R.; McLean, J.P.; Arcangeli, C.; Matthews, R. Dynamic top height models for several major forest tree species in Great Britain. Forestry 2021, 94, 181–192. [Google Scholar] [CrossRef]
- Sharma, R.P.; Brunner, A.; Eid, T.; Øyen, B.H. Modelling dominant height growth from national forest inventory individual tree data with short time series and large age errors. For. Ecol. Manag. 2011, 262, 2162–2175. [Google Scholar] [CrossRef]
- Ercanli, I.; Kahriman, A.; Yavuz, H. Dynamic base-age invariant site index models based on generalized algebraic difference approach for mixed Scots pine (Pinus sylvestris L.) and Oriental beech (Fagus orientalis Lipsky) stands. Turk. J. Agric. For. 2014, 38, 134–147. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S.; Jansa, V.; Kučera, M. Modelling individual tree diameter growth for Norway spruce in the Czech Republic using a generalized algebraic difference approach. J. For. Sci. 2017, 63, 227–238. [Google Scholar] [CrossRef]
- LSFRI. Meža Resursu Monitoringa Metodika (Methodology of Monitoring of Forest Resources). 2022. Available online: https://www.silava.lv/images/Petijumi/Nacionalais-meza-monitorings/2022-04-28-MRM-metodika.pdf (accessed on 1 October 2023).
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Zeide, B. Analysis of growth equations. For. Sci. 1993, 39, 594–616. [Google Scholar] [CrossRef]
- Cieszewski, C.J. Comparing properties of self-referencing models based on nonlinear-fixed-effects versus nonlinear-mixed-effects modeling approaches. Math. Comput. For. Nat.-Resour. Sci. 2018, 10, 46–57. [Google Scholar]
- Nigh, G. Engelmann spruce site index models: A comparison of model functions and parameterisations. PLoS ONE 2015, 10, e0124079. [Google Scholar] [CrossRef] [PubMed]
- Semaškienė, L. Small-Leaved Lime (Tilia cordata Mill.) in Lithuania: Phenotypical Diversity and Productivity of Modal Stands. Ph.D. Thesis, Lithuanian University of Agriculture, Kaunas, Lithuania, 2006. [Google Scholar]
- Böckmann, T. Wachstum und Ertrag der Winterlinde (Tilia cordata Mill.) in Niedersachsen und Nordhessen. Ph.D. Thesis, Universität Göttingen, Göttingen, Germany, 1990. [Google Scholar]
- Socha, J.; Ochał, W.; Grabczyński, S.; Maj, M. Modele bonitacyjne dla gatunków lasotwórczych Polski opracowane na podstawie tablic zasobności. Sylwan 2015, 159, 639–649. [Google Scholar]
- Giurgiu, V.; Drăghiciu, D. Modele Matimatico-Auxologice Şi Tabele De Producţie Pentru Arboreta; Romania Editura Ceres: Bucureşti, Romania, 2004; pp. 537–538. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 15 September 2023).
- Krumland, B.; Eng, H. Site index systems for major young-growth forest woodland species in northern California. Calif. For. 2005, 4, 1–220. [Google Scholar]
- Anta, M.B.; Dieguez-Aranda, U. Site quality of pedunculate oak (Quercus robur L.) stands in Galicia (northwest Spain). Eur. J. For. Res. 2005, 124, 19–28. [Google Scholar] [CrossRef]
- Nagel, J. Wachstumsmodell für Bergahorn in Schleswig-Holstein. Ph.D. Thesis, Universität Göttingen, Göttingen, Germany, 1985. [Google Scholar]
- Kjølby, V. Ær: Naturhistorie, tilvækst og hugst, Ær (Acer pseudoplatanus L.); Dansk Skovforening: København, Denmark, 1958; pp. 5–126. [Google Scholar]
- Pigott, C.D. Tilia cordata miller. J. Ecol. 1991, 79, 1147–1207. [Google Scholar] [CrossRef]
- Socha, J.; Solberg, S.; Tymińska-Czabańska, L.; Tompalski, P.; Vallet, P. Height growth rate of Scots pine in Central Europe increased by 29% between 1900 and 2000 due to changes in site productivity. For. Ecol. Manag. 2021, 490, 119102. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought and waterlogging of temperate, northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Matisons, R.; Šņepsts, G.; Puriņa, L.; Donis, J.; Jansons, Ā. Dominant height growth of European beech at the northeasternmost stands in Europe. Silva Fenn. 2018, 52, 7818. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Tome, M.; Bravo, F.; Montero, G.; del Rio, M. Dominant height growth equations including site attributes in the generalized algebraic difference approach. Can. J. Forest Res. 2008, 38, 2348–2358. [Google Scholar] [CrossRef]
- Sharma, M. Modeling climate effects on site productivity of plantation grown jack pine, black spruce, red pine, and white spruce using annual/seasonal climate values. Forests 2022, 13, 1600. [Google Scholar] [CrossRef]
- Leites, L.P.; Robinson, A.P.; Rehfeldt, G.E.; Marshall, J.D.; Crookston, N.L. Height-growth response to climatic changes differs among populations of Douglas-fir: A novel analysis of historic data. Ecol. Appl. 2012, 22, 154–165. [Google Scholar] [CrossRef] [PubMed]



| Variable (Unit) | Statistic | Small-Leaved Lime | Norway Maple | ||
|---|---|---|---|---|---|
| Calibration | Cross-Validation | Calibration | Cross-Validation | ||
| Age (year) | Mean | 47.5 | 49.2 | 39.0 | 35.7 |
| St. dev. | 22.7 | 23.7 | 24.8 | 19.6 | |
| Min | 3 | 3 | 4 | 4 | |
| Max | 103 | 103 | 122 | 120 | |
| Height (m) | Mean | 19.5 | 19.3 | 17.1 | 17.0 |
| St. dev. | 6.2 | 6.5 | 5.9 | 5.5 | |
| Min | 3.5 | 4.4 | 3.9 | 5.3 | |
| Max | 31.5 | 32.5 | 33.6 | 32.7 | |
| Sample plots (-) | Number | 81 | 47 | 126 | 70 |
| Trees (-) | Number | 197 | 87 | 224 | 92 |
| Measurements of trees (-) | Number | 277 | 100 | 300 | 100 |
| Species | Model | MR (m) | RMSE (m) | adj. R2 | AIC | |
|---|---|---|---|---|---|---|
| Fit statistic | Small-leaved lime (N = 277) | Chapman–Richards | 0.01 | 0.84 | 0.980 | −95.7 |
| Hossfeld | 0.08 | 0.86 | 0.979 | −79.7 | ||
| Hossfeld I | −0.04 | 0.86 | 0.980 | −83.5 | ||
| Hossfeld IV | 0.06 | 0.86 | 0.979 | −80.0 | ||
| Sloboda | 0.01 | 0.84 | 0.980 | −96.2 | ||
| Strand | 0.02 | 0.84 | 0.980 | −97.3 | ||
| Norway maple (N = 300) | Chapman–Richards | 0.01 | 0.94 | 0.984 | −37.1 | |
| Hossfeld | 0.18 | 1.02 | 0.982 | 14.0 | ||
| Hossfeld I | 0.08 | 0.99 | 0.983 | −5.5 | ||
| Hossfeld IV | 0.10 | 1.01 | 0.982 | 8.1 | ||
| Sloboda | 0.01 | 0.94 | 0.984 | −34.5 | ||
| Strand | 0.07 | 0.99 | 0.983 | −6.8 | ||
| Cross-validation | Small-leaved lime (N = 100) | Chapman–Richards | 0.04 | 0.95 | 0.976 | −8.8 |
| Hossfeld | 0.15 | 0.96 | 0.977 | −6.9 | ||
| Hossfeld I | 0.01 | 0.96 | 0.976 | −6.7 | ||
| Hossfeld IV | 0.13 | 0.95 | 0.977 | −7.1 | ||
| Sloboda | 0.05 | 0.94 | 0.976 | −9.3 | ||
| Strand | 0.06 | 0.96 | 0.976 | −5.5 | ||
| Norway maple (N = 100) | Chapman–Richards | −0.10 | 1.04 | 0.981 | 10.5 | |
| Hossfeld | 0.06 | 1.08 | 0.979 | 16.8 | ||
| Hossfeld I | −0.02 | 1.05 | 0.980 | 11.3 | ||
| Hossfeld IV | −0.02 | 1.07 | 0.979 | 16.4 | ||
| Sloboda | −0.10 | 1.04 | 0.981 | 9.6 | ||
| Strand | −0.04 | 1.05 | 0.980 | 12.6 |
| Model | Parameter | Small-Leaved Lime | Norway Maple | ||
|---|---|---|---|---|---|
| Estimate | Standard Error | Estimate | Standard Error | ||
| Chapman–Richards | b1 | 0.0091 | 0.0026 | 0.0254 | 0.0025 |
| b2 | −417.22 | 11.49 | −80.60 | 17.49 | |
| b3 | 1585.68 | 3.01 | 284.27 | 60.35 | |
| Hossfeld | b1 | 63.57 | 10.17 | 50.62 | 7.93 |
| b2 | −44.32 | 0.0111 | −16.2958 | 0.0481 | |
| b3 | 0.8537 | 0.0453 | 1.0402 | 0.0502 | |
| Hossfeld I | b1 | 0.0283 | 0.0011 | 0.0255 | 0.0015 |
| b2 | −0.0061 | 0.0004 | −0.0053 | 0.0005 | |
| Hossfeld IV | b1 | 0.8405 | 0.0469 | 1.0338 | 0.0564 |
| b2 | −1449.54 | 50.22 | −2451.73 | 94.81 | |
| b3 | 970.38 | 75.08 | 1264.81 | 182.70 | |
| Sloboda | b1 | 45.12 | 9.29 | 34.82 | 2.69 |
| b2 | 0.1387 | 0.0212 | 0.1781 | 0.0248 | |
| b3 | 0.5589 | 0.0688 | 0.5024 | 0.0597 | |
| Strand | b1 | 0.0098 | 0.0046 | 0.0342 | 0.0125 |
| b2 | −0.0063 | 0.0005 | −0.0054 | 0.0005 | |
| b3 | 0.7799 | 0.0670 | 1.0769 | 0.1000 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matisone, I.; Šņepsts, G.; Kaupe, D.; Hein, S.; Rieksts-Riekstiņš, R.; Jansons, Ā. Analysis of Height Growth Suggests Moderate Growth of Tilia cordata and Acer platanoides at the Native Hemiboreal Stands in Latvia. Forests 2024, 15, 7. https://doi.org/10.3390/f15010007
Matisone I, Šņepsts G, Kaupe D, Hein S, Rieksts-Riekstiņš R, Jansons Ā. Analysis of Height Growth Suggests Moderate Growth of Tilia cordata and Acer platanoides at the Native Hemiboreal Stands in Latvia. Forests. 2024; 15(1):7. https://doi.org/10.3390/f15010007
Chicago/Turabian StyleMatisone, Ilze, Guntars Šņepsts, Dārta Kaupe, Sebastian Hein, Raitis Rieksts-Riekstiņš, and Āris Jansons. 2024. "Analysis of Height Growth Suggests Moderate Growth of Tilia cordata and Acer platanoides at the Native Hemiboreal Stands in Latvia" Forests 15, no. 1: 7. https://doi.org/10.3390/f15010007